Keynestermit
- anaerobinen: Ilman happea; erityisesti ympäristön tai organismin.
- reduktio: Reaktio, jossa saadaan elektroneja ja pienennetään valenssia; usein poistamalla happea tai lisäämällä vetyä.
- anaerobinen hengitys: Eliöiden soluissa tapahtuvat aineenvaihduntareaktiot ja -prosessit, jotka käyttävät muita elektroniakseptoreita kuin happea
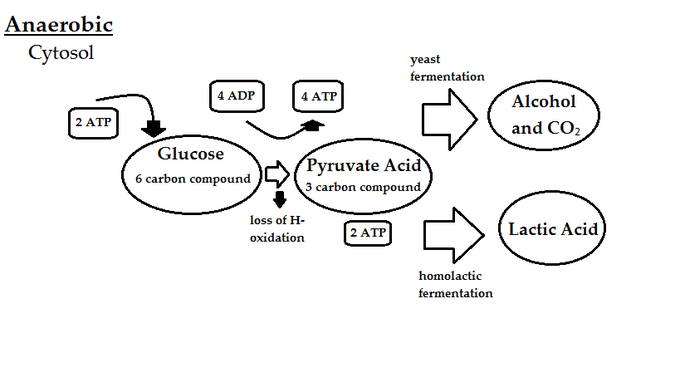
Anaerobinen hengitys on ATP:n muodostumista ilman happea. Tämä menetelmä sisältää edelleen hengityselinten elektroninsiirtoketjun, mutta ilman, että happea käytetään pääteelektroniakseptorina. Sen sijaan elektroniakseptoreina käytetään molekyylejä, kuten sulfaattia (SO42-), nitraattia (NO3-) tai rikkiä (S). Näillä molekyyleillä on alhaisempi pelkistymispotentiaali kuin hapella, joten energiaa muodostuu vähemmän glukoosimolekyyliä kohti anaerobisissa kuin aerobisissa olosuhteissa.
Anaerobisessa hengityksessä voidaan käyttää monia erityyppisiä elektroniakseptoreita. Denitrifikaatio on nitraatin (NO3-) hyödyntämistä pääteelektroniakseptorina. Nitraatilla, kuten hapellakin, on korkea pelkistyspotentiaali. Tämä prosessi on laajalle levinnyt, ja sitä käyttävät monet Proteobakteerien jäsenet. Monet denitrifioivat bakteerit voivat käyttää myös rautaa (Fe3+) ja erilaisia orgaanisia elektroniakseptoreita.
Sulfaattireduktio käyttää elektroniakseptorina sulfaattia (SO2-4) tuottaen metabolisena lopputuotteena rikkivetyä (H2S). Sulfaattireduktio on energeettisesti suhteellisen köyhä prosessi, ja sitä käyttävät monet δ-proteobakteereihin kuuluvat gramnegatiiviset bakteerit. Sitä käyttävät myös grampositiiviset organismit, jotka ovat sukua Desulfotomaculumille tai arkeonille Archaeoglobus.
Sulfaatin pelkistyminen edellyttää elektroninluovuttajien, kuten hiiliyhdisteiden laktaatin ja pyruvaatin (organotrofiset pelkistäjät) tai vetykaasun (litotrofiset pelkistäjät) käyttöä. Jotkut epätavalliset autotrofiset sulfaattia pelkistävät bakteerit, kuten Desulfotignum phosphitoxidans, voivat käyttää elektroninluovuttajana fosfiittia (HPO3-). Toiset, kuten tietyt Desulfovibrio-suvun lajit, kykenevät rikin disproportionointiin (yhden yhdisteen jakaminen elektroninluovuttajaksi ja elektroniakseptoriksi) käyttäen alkuainerikkiä (S0), sulfiittia (SO3-2) ja tiosulfaattia (S2O32-) tuottaakseen sekä rikkivetyä (H2S) että sulfaattia (SO2-).
Acetogeneesi on mikrobien aineenvaihduntatyyppi, joka käyttää vetyä (H2) elektroninluovuttajana ja hiilidioksidia (CO2) elektroniakseptorina tuottaakseen asetaattia, samoja elektroninluovuttajia ja -akseptoreita, joita käytetään metanogeneesissä.
Rauta (Fe3+) on laajalle levinnyt anaerobinen pääte-elektroniakseptori, jota käyttävät sekä autotrofiset että heterotrofiset organismit. Elektronien kulku näissä organismeissa on samanlainen kuin elektronin kuljetuksessa, joka päättyy happiin tai nitraattiin, paitsi että rautarautaa pelkistävissä organismeissa tämän järjestelmän viimeinen entsyymi on rautaraudan reduktaasi. Koska jotkin rautaa pelkistävät bakteerit (esim. G. metallireducens) voivat käyttää myrkyllisiä hiilivetyjä (esim. tolueenia) hiililähteenä, näiden organismien käyttäminen biologisena puhdistusaineena rautarautaa sisältävissä saastuneissa pohjavesissä herättää suurta kiinnostusta.
Muita epäorgaanisia elektronin akseptoreita ovat mangaani-ionin (Mn4+) pelkistyminen mangaaniksi (Mn2+), selenaatin (SeO42-) pelkistyminen seleniitiksi (SeO32-) seleeniksi (Se), arsenaatti (AsO43-) arseniitiksi (AsO33-) ja uraani (UO22+) uraanidioksidiksi (UO2)
Orgaanisia yhdisteitä voidaan käyttää myös elektronien akseptoreina anaerobisessa hengityksessä. Tällaisia ovat esimerkiksi fumaraatin pelkistyminen sukkinaatiksi, trimetyyliamiini-N-oksidin (TMAO) pelkistyminen trimetyyliamiiniksi (TMA) ja dimetyylisulfoksidin (DMSO) pelkistyminen dimetyylisulfidiksi (DMS).